
Resolution of the Hypoxylon fuscum Complex (Hypoxylaceae, Xylariales) and Discovery and Biological Characterization of Two of Its Prominent Secondary Metabolites

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Sources
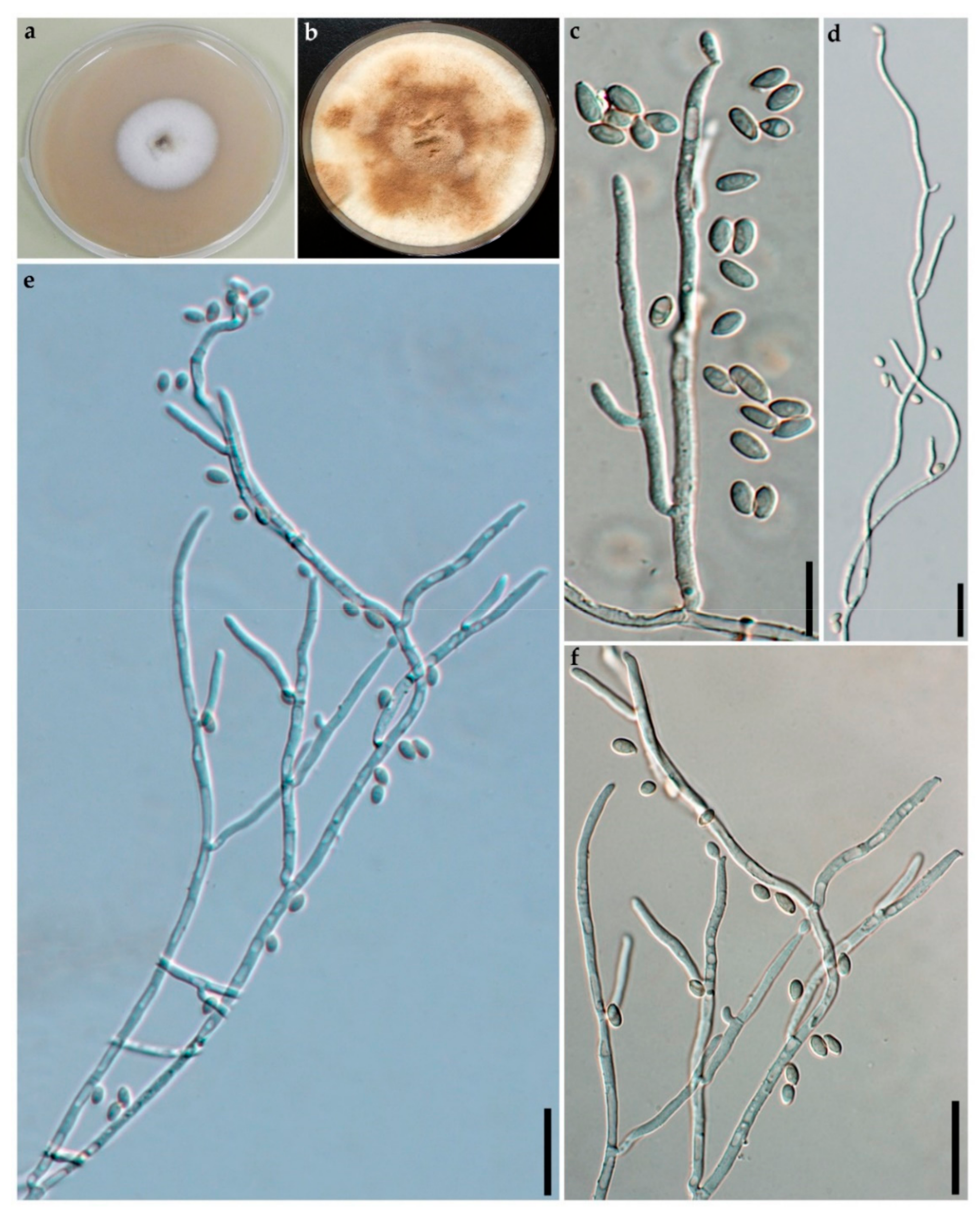
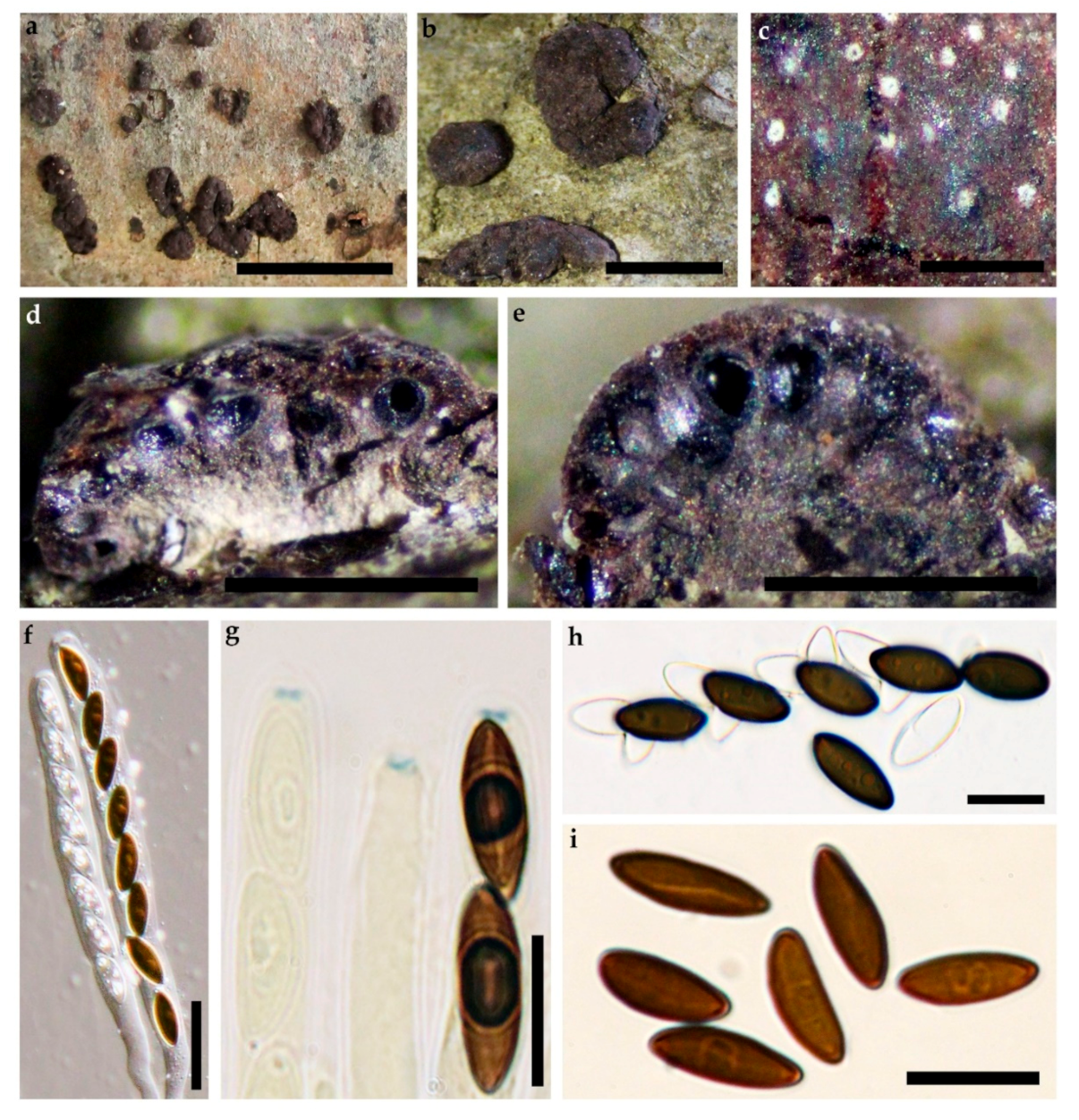
2.2. Morphological Characterisation
2.3. DNA Extraction, PCR and Sequencing
2.4. Molecular Phylogenetic Analyses
2.5. HPLC Profiling
2.6. Extraction and Isolation of Compounds 4, 5, and 6
2.7. Spectral Data
2.7.1. Pseudofuscochalasin A (4) Figures S1–S6
2.7.2. Daldinin F (5) Figures S7–S11
2.7.3. 3′-Malonyl-daldinin F (6) Figures S12–S17
2.8. Antimicrobial Activities and Cytotoxicity
2.9. Actin Disruption Assay
3. Results
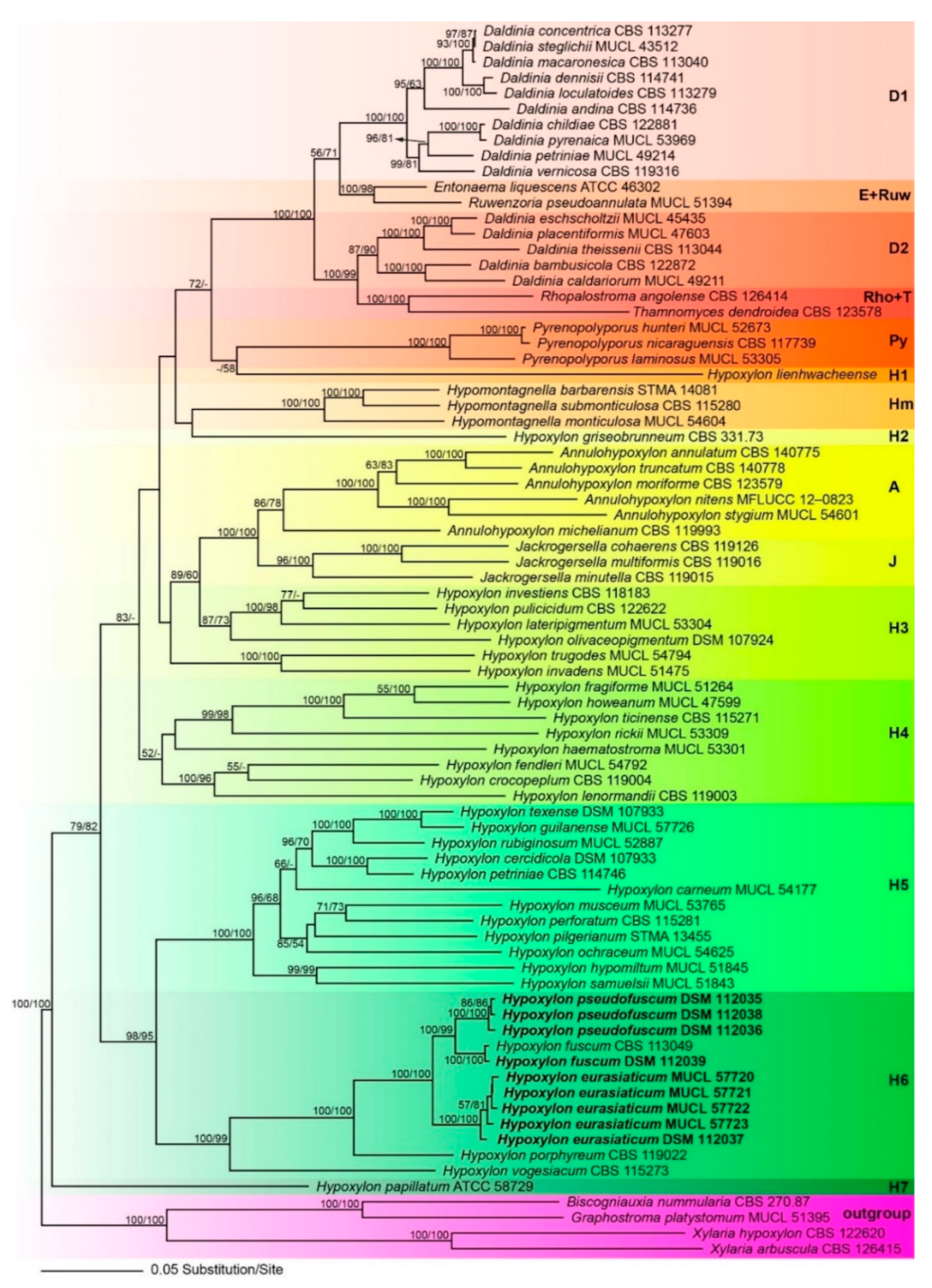
3.1. Molecular Phylogeny
3.2. Taxonomic Part
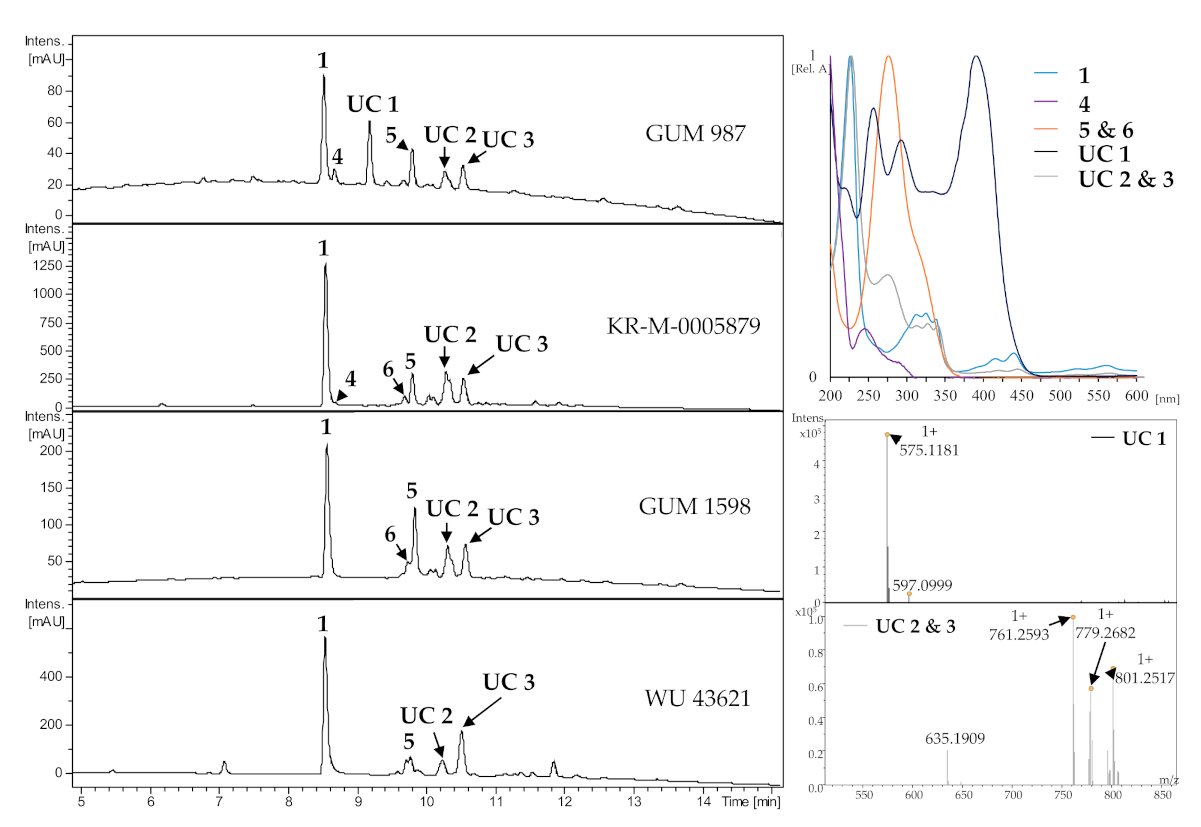
3.3. HPLC Profiling
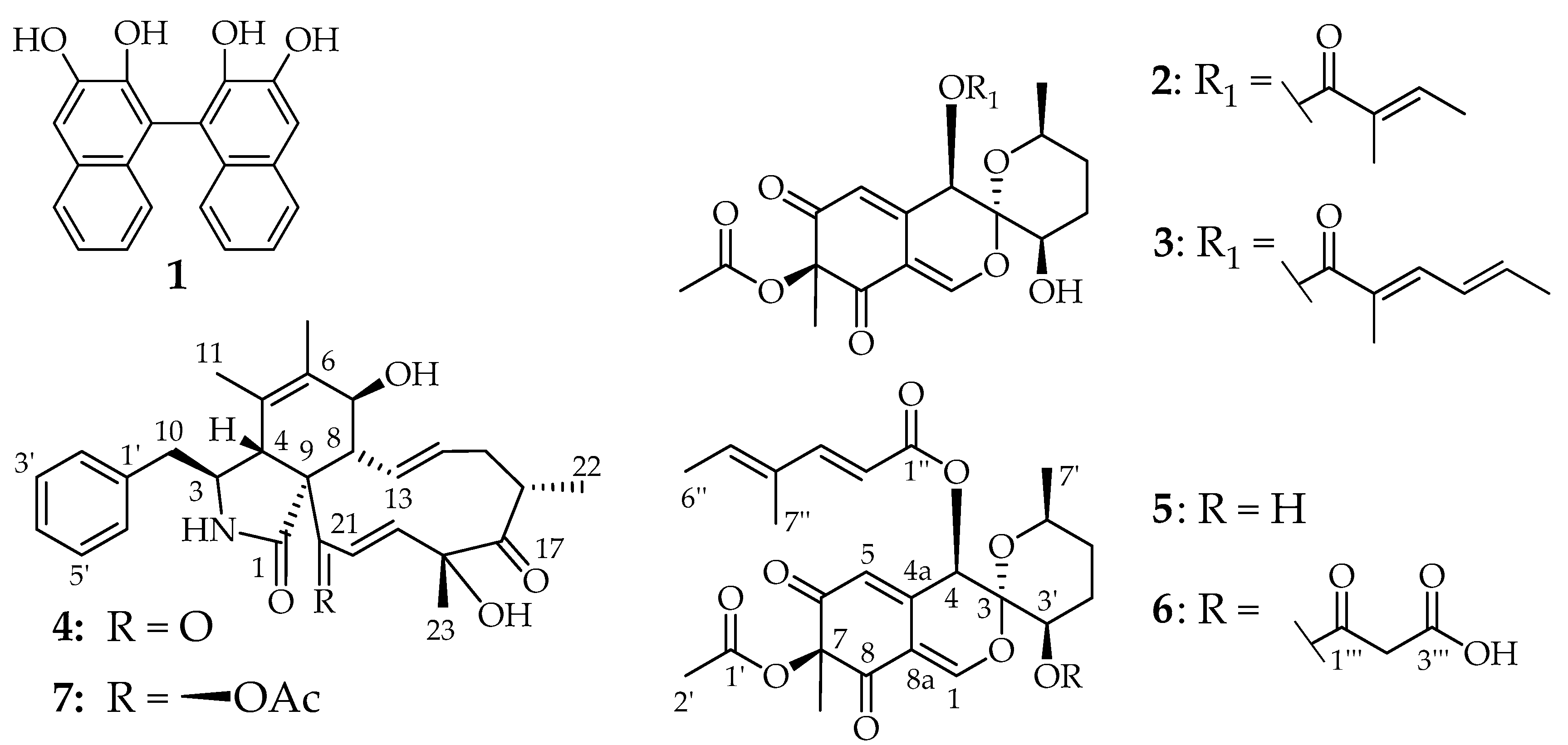
3.4. Structure Elucidation (Figure 5 and Figure 6)
3.5. Antibacterial and Cytotoxic Activities
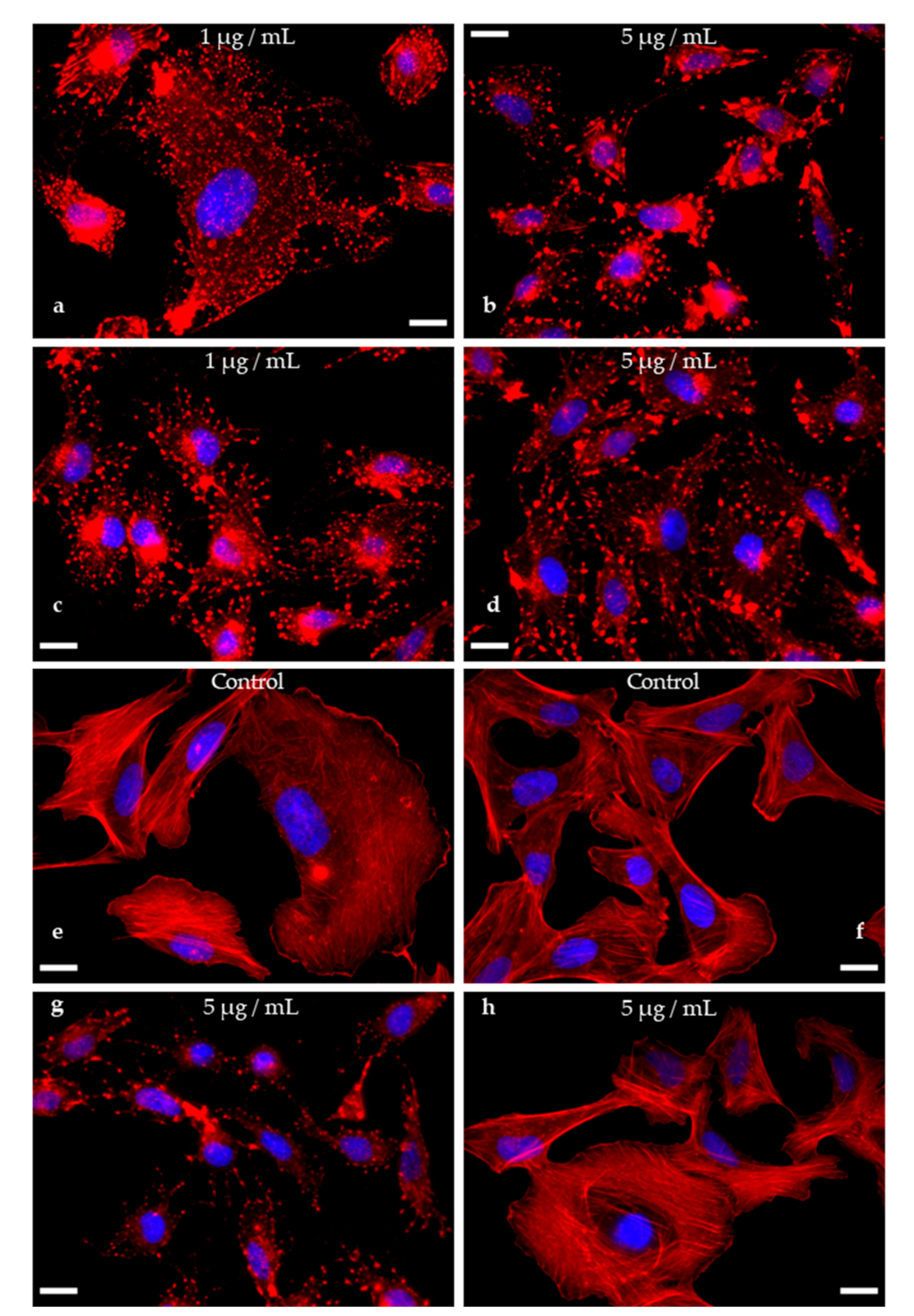
3.6. Actin Disruption Assay (Figure 7)
4. Discussion
5. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.; Bhat, D.J.; Jones, E.B.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef] [Green Version]
- Pažoutová, S.; Follert, S.; Bitzer, J.; Keck, M.; Surup, F.; Šrůtka, P.; Holuša, J.; Stadler, M. A new endophytic insect-associated Daldinia species, recognised from a comparison of secondary metabolite profiles and molecular phylogeny. Fungal Divers. 2013, 60, 107–123. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D. A Revision of the Genus Hypoxylon; Mycologia Memoir n° 20; APS Press: St. Paul, MN, USA, 1996; p. 365. [Google Scholar]
- Daranagama, D.A.; Hyde, K.D.; Sir, E.B.; Thambugala, K.M.; Tian, Q.; Samarakoon, M.C.; McKen-zie, E.H.C.; Jayasiri, S.C.; Tibpromma, S.; Bhat, J.D.; et al. Towards a natural classification and back-bone tree for Graphostromataceae, Hypoxylaceae, Lopadostomataceae and Xylariaceae. Fungal Divers. 2018, 88, 1–165. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Wendt, L.; Hladki, A.I.; Romero, A.I.; Stadler, M.; Sir, E.B. Hypomontagnella (Hypoxylaceae): A new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol. Prog. 2019, 18, 187–201. [Google Scholar] [CrossRef]
- Wibberg, D.; Stadler, M.; Lambert, C.; Bunk, B.; Spröer, C.; Rückert, C.; Kalinowski, J.; Cox, R.J.; Kuhnert, E. High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers. 2020. [Google Scholar] [CrossRef]
- Granmo, A. Morphotaxonomy and Chorology of the Genus Hypoxylon (Xylariaceae) in Norway. Ph.D. Thesis, Botanical Garden and Museum-University of Oslo, Oslo, Norway, 1999. [Google Scholar]
- Stadler, M.; Fournier, J.; Granmo, A.; Beltrán-Tejera, E. The “red Hypoxylons” of the temperate and subtropical Northern Hemisphere. N. Am. Fungi 2008, 3, 73–125. [Google Scholar] [CrossRef] [Green Version]
- Fournier, J.; Köpcke, B.; Stadler, M. New species of Hypoxylon from Western Europe and Ethiopia. Mycotaxon 2010, 113, 209–235. [Google Scholar] [CrossRef]
- Quang, D.N.; Hashimoto, T.; Tanaka, M.; Stadler, M.; Asakawa, Y. Cyclic azaphilones daldinins E and F from the ascomycete fungus Hypoxylon fuscum (Xylariaceae). Phytochemistry 2004, 65, 469–473. [Google Scholar] [CrossRef]
- Petrini, L.E. Haupt-und Nebenfruchtformen europäischer Hypoxylon-Arten (Xylariaceae, Sphaeriales) und verwandter Pilze. Mycol. Helv. 1986, 1, 501–627. [Google Scholar]
- Petrini, L.E.; Petrini, O.; Sieber, T.N. Host specificity of Hypoxylon fuscum: A statistical approach to the problem. Sydowia 1987, 40, 227–234. [Google Scholar]
- Stadler, M.; Fournier, J. Pigment chemistry, taxonomy and phylogeny of the Hypoxyloideae (Xylariaceae). Rev. Iberoam. Micol. 2006, 23, 160–170. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Quang, D.N.; Akulov, A.Y. Metabolomic studies on the chemical ecology of the Xylariaceae (Ascomycota). Nat. Prod. Commun. 2007, 2, 287–304. [Google Scholar] [CrossRef] [Green Version]
- Triebel, D.; Peršoh, D.; Wollweber, H.; Stadler, M. Phylogenetic relationships among Daldinia, Entonaema and Hypoxylon as inferred from ITS nrDNA sequences. Nova Hedwig. 2005, 80, 25–43. [Google Scholar] [CrossRef]
- Raei, S.; Khodaparast, S.A.; Abbasi, M. Contribution to the knowledge of Hypoxylon and Annulohypoxylon in Guilan province (N Iran). Rostaniha 2012, 13, 197–206. [Google Scholar] [CrossRef]
- Raei, S.; Khodaparast, S.A.; Abbasi, M. More records of xylariaceous fungi from North of Iran. Rostaniha 2014, 15, 110–121. [Google Scholar] [CrossRef]
- Pourmoghaddam, M.J.; Khodaparast, S.A.; Pedramfar, H. The genus Daldinia in Guilan province (N Iran). Rostaniha 2014, 15, 122–132. [Google Scholar] [CrossRef]
- Pourmoghaddam, M.J.; Lambert, C.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stadler, M. Discovery of a new species of the Hypoxylon rubiginosum complex from Iran and antagonistic activities of Hypoxylon spp. against the Ash Dieback pathogen, Hymenoscyphus fraxineus, in dual culture. MycoKeys 2020, 66, 105–133. [Google Scholar] [CrossRef] [Green Version]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute (Great Britain); British Mycological Society: Manchester, UK, 1970. [Google Scholar]
- Kuhnert, E.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Rohde, M.; Stadler, M. Phylogenetic and chemotaxonomic resolution of the genus Annulohypoxylon (Xylariaceae) including four new species. Fungal Divers. 2017, 85, 1–43. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. In Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP* 4.0b10: Phylogenetic Analysis Using Parsimony (*and other Methods); Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Peršoh, D.; Luangsa-ard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Bitzer, J.; Laessoe, T.; Fournier, J.; Kummer, V.; Decock, C.; Tichy, H.V.; Piepenbring, M.; Persoh, D.; Stadler, M. Affinities of Phylacia and the daldinoid Xylariaceae, inferred from chemotypes of cultures and ribosomal DNA sequences. Mycol. Res. 2008, 112, 251–270. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American endophytes to the phylogeny, ecology, and taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenetics Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groe-newald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Stadler, M.; Laessoe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Persoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Castlebury, L.A.; Miller, A.N.; Huhndorf, S.M.; Schoch, C.L.; Seifert, K.A.; Rossman, A.Y.; Rogers, J.D.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; et al. An overview of the systematics of the Sordariomycetes based on a four-gene phylogeny. Mycologia 2006, 98, 1076–1087. [Google Scholar] [CrossRef]
- Koukol, O.; Kelnarová, I.; Černý, K. Recent observations of sooty bark disease of sycamore maple in Prague (Czech Republic) and the phylogenetic placement of Cryptostroma corticale. For. Pathol. 2015, 45, 21–27. [Google Scholar] [CrossRef]
- Stadler, M.; Kuhnert, E.; Peršoh, D.; Fournier, J. The Xylariaceae as model example for a unified nomenclature following the “One Fungus-One Name” (1F1N) concept. Mycology 2013, 4, 5–21. [Google Scholar] [CrossRef]
- Becker, K.; Lambert, C.; Wieschhaus, J.; Stadler, M. Phylogenetic assignment of the fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and investigation of its secondary metabolites. Microorganisms 2020, 8, 1397. [Google Scholar] [CrossRef] [PubMed]
- Sir, E.B.; Becker, K.; Lambert, C.; Bills, F.G.; Kuhnert, E. Observations on Texas hypoxylons, including two new Hypoxylon species and widespread environmental isolates of the H. croceum complex identified by a polyphasic approach. Mycologia 2019, 111, 832–856. [Google Scholar] [CrossRef] [PubMed]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Bills, G.F.; Gonzalez-Menendez, V.; Platas, G.; Fournier, J.; Persoh, D.; Stadler, M. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 2012, e46687. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Læssøe, T.; Chlebicki, A.; Lechat, C.; Flessa, F.; Rambold, G.; Peršoh, D. Chemotaxonomic and phylogenetic studies of Thamnomyces (Xylariaceae). Mycoscience 2010, 51, 189–207. [Google Scholar] [CrossRef]
- Fournier, J.; Flessa, F.; Peršoh, D.; Stadler, M. Three new Xylaria species from southwestern Europe. Mycol. Prog. 2011, 10, 33–52. [Google Scholar] [CrossRef]
- Sir, E.B.; Kuhnert, E.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Stadler, M. New species and reports of Hypoxylon from Argentina recognized by a polyphasic approach. Mycol. Prog. 2016, 15, 42. [Google Scholar] [CrossRef]
- Moussa, A.Y.; Lambert, C.; Stradal, T.E.; Ashrafi, S.; Maier, W.; Stadler, M.; Helaly, S.E. New peptaibiotics and a cyclodepsipeptide from Ijuhya vitellina: Isolation, identification, cytotoxic and nematicidal activities. Antibiotics 2020, 9, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandargo, B.; Michehl, M.; Stadler, M.; Surup, F. Antifungal sesquiterpenoids, rhodocoranes F-L from submerged cultures of the wrinkled peach mushroom, Rhodotus palmatus. J. Nat. Prod. 2020, 83, 720–724. [Google Scholar] [CrossRef]
- Kretz, R.; Wendt, L.; Wongkanoun, S.; Luangsa-ard, J.J.; Surup, F.; Helaly, S.E.; Noumeur, S.R.; Stadler, M.; Stradal, T.E. The effect of cytochalasans on the actin cytoskeleton of eukaryotic cells and preliminary structure–activity relationships. Biomolecules 2019, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, M.S.; Hashimoto, T.; Asakawa, Y. Five 10-phenyl-[11]-cytochalasans from a Daldinia fungal species. Phytochemistry 1995, 40, 135–140. [Google Scholar] [CrossRef]
- Suzuki, S.; Hosoe, T.; Nozawa, K.; Yaguchi, T.; Udagawa, S.; Kawai, K. Mitorubrin derivatives on ascomata of some Talaromyces species of ascomycetous fungi. J. Nat. Prod. 1999, 62, 1328–1329. [Google Scholar] [CrossRef]
- Clark, R.C.; Lee, S.Y.; Boger, D.L. Total synthesis of chlorofusin, its seven chromophore diastereomers, and key partial structures. J. Am. Chem. Soc. 2008, 130, 12355–12369. [Google Scholar] [CrossRef] [Green Version]
- Yahara, I.; Harada, F.; Sekita, S.; Yoshihira, K.; Natori, S. Correlation between effects of 24 different cytochalasins on cellular structures and cellular events and those on actin in vitro. J. Cell Biol. 1982, 92, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Mühlbauer, A.; Triebel, D.; Persoh, D.; Wollweber, H.; Seip, S.; Stadler, M. Macrocarpones, novel metabolites from stromata of Hypoxylon macrocarpum and new evidence on the chemotaxonomy of Hypoxylon species. Mycol. Prog. 2020, 1, 235–248. [Google Scholar] [CrossRef]
- Hongsanan, S.; Xie, N.; Liu, J.K.; Dissanayake, A.; Ekanayaka, A.H.; Raspé, O.; Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Purahong, W.; et al. Can we use environmental DNA as holotypes? Fungal Divers. 2018, 92, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Stadler, M.; Lambert, C.; Wibberg, D.; Kalinowski, J.; Cox, R.J.; Kolařík, M.; Kuhnert, E. Intragenomic polymorphisms in the ITS region of high-quality genomes of the Hypoxylaceae (Xylariales, Ascomycota). Mycol. Prog. 2020, 19, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Quang, D.N.; Hashimoto, T.; Radulovic, N.; Stadler, M.; Asakawa, Y. Antimicrobial azaphilones from the xylariaceous inedible mushrooms. Int. J. Med. Mushrooms 2005, 7, 452–455. [Google Scholar] [CrossRef]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Nat. Prod. Rep. 2010, 27, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Basnet, B.B.; Chen, B.; Suleimen, Y.M.; Ma, K.; Guo, S.; Bao, L.; Huang, Y.; Liu, H. Cytotoxic secondary metabolites from the endolichenic fungus Hypoxylon fuscum. Planta Med. 2019, 13, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Widmer, T.L.; McMahon, M.B.; Luster, D.G. Plant pathogenic fungi are harbored as endophytes in Rhododendron spp. native to the Eastern USA. Fungal Ecol. 2020, 47, 100949. [Google Scholar] [CrossRef]
- Becker, K.; Stadler, M. Recent progress in biodiversity research on the Xylariales and their secondary metabolism. J. Antibiot. 2021, 74, 1–23. [Google Scholar] [CrossRef]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number | Origin | Status | GenBank Accession Number | References | |||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | RPB2 | TUB2 | |||||
| Annulohypoxylon annulatum | CBS 140775 | Texas | ET | KY610418 | KY610418 | KY624263 | KX376353 | [2,23] |
| Annulohypoxylon michelianum | CBS 119993 | Spain | KX376320 | KY610423 | KY624234 | KX271239 | [2,29] | |
| Annulohypoxylon moriforme | CBS 123579 | Martinique | KX376321 | KY610425 | KY624289 | KX271261 | [2,23] | |
| Annulohypoxylon nitens | MFLUCC 12–0823 | Thailand | KJ934991 | KJ934992 | KJ934994 | KJ934993 | [30] | |
| Annulohypoxylon stygium | MUCL 54601 | French Guiana | KY610409 | KY610475 | KY624292 | KX271263 | [2] | |
| Annulohypoxylon truncatum | CBS 140778 | Texas | ET | KY610419 | KY610419 | KY624277 | KX376352 | [2,23] |
| Biscogniauxia nummularia | MUCL 51395 | France | ET | KY610382 | KY610427 | KY624236 | KX271241 | [2] |
| Daldinia andina | CBS 114736 | Ecuador | HT | AM749918 | KY610430 | KY624239 | KC977259 | [2,29,31] |
| Daldinia bambusicola | CBS 122872 | Thailand | HT | KY610385 | KY610431 | KY624241 | AY951688 | [2,6] |
| Daldinia caldariorum | MUCL 49211 | France | AM749934 | KY610433 | KY624242 | KC977282 | [2,29,31] | |
| Daldinia childiae | CBS 122881 | France | HT | KU683757 | MH874773 | KU684290 | KU684129 | [32,33] |
| Daldinia concentrica | CBS 113277 | Germany | AY616683 | KY610434 | KY624243 | KC977274 | [2,17,29] | |
| Daldinia dennisii | CBS 114741 | Australia | HT | JX658477 | KY610435 | KY624244 | KC977262 | [2,29,34] |
| Daldinia eschscholtzii | MUCL 45435 | Benin | JX658484 | KY610437 | KY624246 | KC977266 | [2,29,34] | |
| Daldinia loculatoides | CBS 113279 | UK | ET | AF176982 | KY610438 | KY624247 | KX271246 | [2] |
| Daldinia macaronesica | CBS 113040 | Spain | PT | KY610398 | KY610477 | KY624294 | KX271266 | [2] |
| Daldinia petriniae | MUCL 49214 | Austria | ET | AM749937 | KY610439 | KY624248 | KC977261 | [2,29,31] |
| Daldinia placentiformis | MUCL 47603 | Mexico | AM749921 | KY610440 | KY624249 | KC977278 | [2,29,31] | |
| Daldinia pyrenaica | MUCL 53969 | France | KY610413 | KY610413 | KY624274 | KY624312 | [2] | |
| Daldinia steglichii | MUCL 43512 | Papua New Guinea | PT | KY610399 | KY610479 | KY624250 | KX271269 | [2] |
| Daldinia theissenii | CBS 113044 | Argentina | PT | KY610388 | KY610441 | KY624251 | KX271247 | [2] |
| Daldinia vernicosa | CBS 119316 | Germany | ET | KY610395 | KY610442 | KY624252 | KC977260 | [2,29] |
| Entonaema liquescens | ATCC 46302 | USA | KY610389 | KY610443 | KY624253 | KX271248 | [2] | |
| Graphostroma platystomum | CBS 270.87 | France | JX658535 | DQ836906 | KY624296 | HG934108 | [2,34,35,36] | |
| Hypomontagnella barbarensis | STMA 14081 | Argentina | HT | MK131720 | MK131718 | MK135891 | MK135893 | [7] |
| Hypomontagnella monticulosa | MUCL 54604 | French Guiana | ET | KY610404 | KY610487 | KY624305 | KX271273 | [2] |
| Hypomontagnella submonticulosa | CBS 115280 | France | KC968923 | KY610457 | KY624226 | KC977267 | [2,29] | |
| Hypoxylon carneum | MUCL 54177 | France | KY610400 | KY610480 | KY624297 | KX271270 | [2] | |
| Hypoxylon cercidicola | CBS 119009 | France | KC968908 | KY610444 | KY624254 | KC977263 | [2,29] | |
| Hypoxylon crocopeplum | CBS 119004 | France | KC968907 | KY610445 | KY624255 | KC977268 | [2,29] | |
| Hypoxylon eurasiaticum | MUCL 57720 | Iran | HT | MW367851 | not obtained | MW373852 | MW373861 | This study |
| Hypoxylon eurasiaticum | MUCL 57721 | Iran | MW367852 | not obtained | MW373853 | MW373862 | This study | |
| Hypoxylon eurasiaticum | MUCL 57722 | Iran | MW367853 | not obtained | MW373854 | MW373863 | This study | |
| Hypoxylon eurasiaticum | MUCL 57723 | Iran | MW367854 | not obtained | MW373855 | MW373864 | This study | |
| Hypoxylon eurasiaticum | DSM 112037 | Poland | MW367855 | not obtained | MW373856 | MW373865 | This study | |
| Hypoxylon fendleri | MUCL 54792 | French Guiana | KF234421 | KY610481 | KY624298 | KF300547 | [2,29] | |
| Hypoxylon fragiforme | MUCL 51264 | Germany | ET | KC477229 | KM186295 | KM186296 | KX271282 | [2,30,37] |
| Hypoxylon fuscum sensu stricto | CBS 113049 | France | ET | KY610401 | KY610482 | KY624299 | KX271271 | [2] |
| Hypoxylon fuscum s. str. | DSM 112039 | Austria | MW367856 | MW367847 | MW373857 | MW373866 | This study | |
| Hypoxylon griseobrunneum | CBS 331.73 | India | HT | KY610402 | KY610483 | KY624300 | KC977303 | [2,29] |
| Hypoxylon guilanense | MUCL 57726 | Iran | HT | MT214997 | MT214992 | MT212235 | MT212239 | [21] |
| Hypoxylon haematostroma | MUCL 53301 | Martinique | ET | KC968911 | KY610484 | KY624301 | KC977291 | [2,29] |
| Hypoxylon howeanum | MUCL 47599 | Germany | AM749928 | KY610448 | KY624258 | KC977277 | [2,29,31] | |
| Hypoxylon hypomiltum | MUCL 51845 | Guadeloupe | KY610403 | KY610449 | KY624302 | KX271249 | [2] | |
| Hypoxylon invadens | MUCL 51475 | France | HT | MT809133 | MT809132 | MT813037 | MT813038 | [38] |
| Hypoxylon investiens | CBS 118183 | Malaysia | KC968925 | KY610450 | KY624259 | KC977270 | [2,29] | |
| Hypoxylon lateripigmentum | MUCL 53304 | Martinique | HT | KC968933 | KY610486 | KY624304 | KC977290 | [2,29] |
| Hypoxylon lenormandii | CBS 119003 | Ecuador | KC968943 | KY610452 | KY624261 | KC977273 | [2,29] | |
| Hypoxylon ochraceum | MUCL 54625 | Martinique | ET | KC968937 | N/A | KY624271 | KC977300 | [2,29] |
| Hypoxylon lienhwacheense | MFLUCC 14-1231 | Thailand | KU604558 | MK287550 | MK287563 | KU159522 | [39,40] | |
| Hypoxylon musceum | MUCL 53765 | Guadeloupe | KC968926 | KY610488 | KY624306 | KC977280 | [2,29] | |
| Hypoxylon olivaceopigmentum | DSM 107924 | USA | HT | MK287530 | MK287542 | MK287555 | MK287568 | [39] |
| Hypoxylon papillatum | ATCC 58729 | USA | HT | KC968919 | KY610454 | KY624223 | KC977258 | [2,29] |
| Hypoxylon perforatum | CBS 115281 | France | KY610391 | KY610455 | KY624224 | KX271250 | [2] | |
| Hypoxylon petriniae | CBS 114746 | France | HT | KY610405 | KY610491 | KY624279 | KX271274 | [2,23] |
| Hypoxylon pilgerianum | STMA 13455 | Martinique | KY610412 | KY610412 | KY624308 | KY624315 | [2] | |
| Hypoxylon porphyreum | CBS 119022 | France | KC968921 | KY610456 | KY624225 | KC977264 | [2,29] | |
| Hypoxylon pseudofuscum | DSM 112038 | Germany | HT | MW367857 | MW367848 | MW373858 | MW373867 | This study |
| Hypoxylon pseudofuscum | DSM 112035 | Germany | MW367858 | MW367849 | MW373859 | MW373868 | This study | |
| Hypoxylon pseudofuscum | DSM 112036 | Germany | MW367859 | MW367850 | MW373860 | MW373869 | This study | |
| Hypoxylon pulicicidum | CBS 122622 | Martinique | HT | JX183075 | KY610492 | KY624280 | JX183072 | [2,41] |
| Hypoxylon rickii | MUCL 53309 | Martinique | ET | KC968932 | KY610416 | KY624281 | KC977288 | [2,29] |
| Hypoxylon rubiginosum | MUCL 52887 | Germany | ET | KC477232 | KY610469 | KY624266 | KY624311 | [2,29] |
| Hypoxylon samuelsii | MUCL 51843 | Guadeloupe | ET | KC968916 | KY610466 | KY624269 | KC977286 | [2,29] |
| Hypoxylon texense | DSM 107933 | USA | HT | MK287536 | MK287548 | MK287561 | MK287574 | [39] |
| Hypoxylon ticinense | CBS 115271 | France | JQ009317 | KY610471 | KY624272 | AY951757 | [2,6] | |
| Hypoxylon trugodes | MUCL 54794 | Sri Lanka | ET | KF234422 | KY610493 | KY624282 | KF300548 | [2,29] |
| Hypoxylon vogesiacum | CBS 115273 | France | KC968920 | KY610417 | KY624283 | KX271275 | [2,23,29] | |
| Jackrogersella cohaerens | CBS 119126 | Germany | KY610396 | KY610497 | KY624270 | KY624314 | [2] | |
| Jackrogersella minutella | CBS 119015 | Portugal | KY610381 | KY610424 | KY624235 | KX271240 | [2,23] | |
| Jackrogersella multiformis | CBS 119016 | Germany | ET | KC477234 | KY610473 | KY624290 | KX271262 | [2,23,29] |
| Pyrenopolyporus hunteri | MUCL 52673 | Ivory Coast | ET | KY610421 | KY610472 | KY624309 | KU159530 | [2,23] |
| Pyrenopolyporus laminosus | MUCL 53305 | Martinique | HT | KC968934 | KY610485 | KY624303 | KC977292 | [2,29] |
| Pyrenopolyporus nicaraguensis | CBS 117739 | Burkina_Faso | AM749922 | KY610489 | KY624307 | KC977272 | [2,29,31] | |
| Rhopalostroma angolense | CBS 126414 | Ivory Coast | KY610420 | KY610459 | KY624228 | KX271277 | [2] | |
| Ruwenzoria pseudoannulata | MUCL 51394 | D. R. Congo | HT | KY610406 | KY610494 | KY624286 | KX271278 | [2] |
| Thamnomyces dendroidea | CBS 123578 | French Guiana | HT | FN428831 | KY610467 | KY624232 | KY624313 | [2,42] |
| Xylaria arbuscula | CBS 126415 | Germany | KY610394 | KY610463 | KY624287 | KX271257 | [2,43] | |
| Xylaria hypoxylon | CBS 122620 | Sweden | ET | KY610407 | KY610495 | KY624231 | KX271279 | [2,44] |
| Taxon | Designation No/(Status) | Ascospores (µm) | Mean (µm) | Host | Known Distribution | KOH-Extractable Pigments | Secondary Metabolites |
|---|---|---|---|---|---|---|---|
| Hypoxylon fuscum sensu stricto | Ww3723/Epitype | 12.5–15.5 × 5–7 | 13.2 × 5.8 | Corylus avellana | Europe | Amber (47) to Honey (64) | 1,5 |
| Hypoxylon fuscum sensu stricto | WU 43621 | 13–15.8× 4.8–6 | 14.4 × 5.4 | Corylus avellana | Austria | Amber (47) to Honey (64) | 1,5 |
| Hypoxylon eurasiaticum | GUM 1597 (H) | 10–12.5 × 4.5–6 | 11.25 × 5.25 | Quercus castaneifolia | Iran | Amber (47) to Orange (7) | 1,5,6 |
| Hypoxylon eurasiaticum | KR-M-0005886 | 9–12 × 3.8–5 | 10.5 × 4.4 | cf. Betula | Poland | Isabelline (65), Olivaceous (47) or Hazel (88) | 1,5,6 |
| Hypoxylon pseudofuscum | KR-M-0005879 (H) | 12–16 × 4.8–7.3 | 14 × 6 | Alnus glutinosa | Germany | Isabelline (65), or Hazel (88) | 1,4,5,6 |
| Hypoxylon pseudofuscum | GUM 987 | 11–15 × 4.5–6.5 | 13 × 5.5 | Alnus sp. | Iran | Amber (47) to Oramge (7) | 1,4,5,6 |
| Hypoxylon pseudofuscum | KR-M-0005877/ KR-M-0005876 | 11–15 × 5.5–6.5 | 13 × 6 | Salix sp. | Germany | Isabelline (65), Olivaceous (47) or Hazel (88) | 1,4,5,6 |
| Species | Specimen | Plant Host | Origin | 1 | 4 | 5 | 6 | UC 1 | UC 2 | UC 3 |
|---|---|---|---|---|---|---|---|---|---|---|
| Hypoxylon fuscum sensu stricto | WU 43621 | Corylus avellana | Austria | + | − | + | − | − | + | + |
| Hypoxylon eurasiaticum | GUM 1597 (H) | Quercus castaneifolia | Iran | + | − | + | + | − | + | + |
| Hypoxylon eurasiaticum | GUM 1598 | Quercus castaneifolia | Iran | + | − | + | + | − | + | + |
| Hypoxylon eurasiaticum | GUM 1600 | Quercus castaneifolia | Iran | + | − | + | + | − | + | + |
| Hypoxylon eurasiaticum | GUM 988 | Quercus castaneifolia | Iran | + | − | + | + | − | + | + |
| Hypoxylon eurasiaticum | KR-M-0005886 | cf. Betula | Poland | + | − | + | + | − | + | + |
| Hypoxylon pseudofuscum | KR-M-0005879 (H) | Alnus glutinosa | Germany | + | + | + | + | + | + | + |
| Hypoxylon pseudofuscum | GUM 987 | Alnus sp. | Iran | + | + | + | + | + | + | + |
| Hypoxylon pseudofuscum | KR-M-0005877 | Salix sp. | Germany | + | + | + | + | + | + | + |
| Hypoxylon pseudofuscum | KR-M-0005876 | Salix sp. | Germany | + | + | + | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambert, C.; Pourmoghaddam, M.J.; Cedeño-Sanchez, M.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stradal, T.E.B.; Stadler, M. Resolution of the Hypoxylon fuscum Complex (Hypoxylaceae, Xylariales) and Discovery and Biological Characterization of Two of Its Prominent Secondary Metabolites. J. Fungi 2021, 7, 131. https://doi.org/10.3390/jof7020131
Lambert C, Pourmoghaddam MJ, Cedeño-Sanchez M, Surup F, Khodaparast SA, Krisai-Greilhuber I, Voglmayr H, Stradal TEB, Stadler M. Resolution of the Hypoxylon fuscum Complex (Hypoxylaceae, Xylariales) and Discovery and Biological Characterization of Two of Its Prominent Secondary Metabolites. Journal of Fungi. 2021; 7(2):131. https://doi.org/10.3390/jof7020131
Chicago/Turabian StyleLambert, Christopher, Mohammad Javad Pourmoghaddam, Marjorie Cedeño-Sanchez, Frank Surup, Seyed Akbar Khodaparast, Irmgard Krisai-Greilhuber, Hermann Voglmayr, Theresia E. B. Stradal, and Marc Stadler. 2021. "Resolution of the Hypoxylon fuscum Complex (Hypoxylaceae, Xylariales) and Discovery and Biological Characterization of Two of Its Prominent Secondary Metabolites" Journal of Fungi 7, no. 2: 131. https://doi.org/10.3390/jof7020131